真核生物の転写調節【遺伝子発現】高校生物発展
概要
動画投稿日|2021年5月17日
動画の長さ|22:39
【 note : https://note.com/yaguchihappy 】
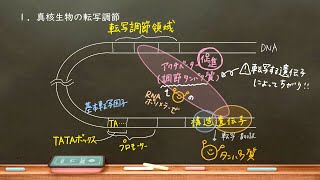
真核生物の遺伝子発現調節(転写調節機構)について講義します。大学内容まで半歩足を踏み入れます。
語呂「基本はプロにもあるぜ。(基本転写因子は、プロモーターにRNAポリメラーゼが結合するのを助ける。あるぜ、がRNAポリメラーゼを表す)」
問題:以下の空欄を埋めよ。
「真核細胞では、( ① )とプロモーターが正しく結合するためには( ② )という一群のタンパク質が必要である。」
答え:①RNAポリメラーゼ ②基本転写因子
●実際には、活性化因子と基本転写因子、RNAポリメラーゼの間で、別のタンパク質が仲立ちをして、間接的に相互作用が行われることが多い。(多くの場合転写因子はDNA非結合性のメディエーターというタンパク質を介して基本転写因子やRNAポリメラーゼに作用する)
●原核生物のσ因子は、RNAポリメラーゼのサブユニットの一つとすることが多い。
●遺伝子発現の主要な調節の段階は転写であると考えられているが、転写調節(今回の動画で紹介したような転写段階の調節)だけでなく、翻訳調節(翻訳の段階での調節)もある。
●サイレンサーに転写抑制因子(リプレッサータンパク質とよぶこともある)が、エンハンサーに転写活性化因子(アクチベータータンパク質とよぶこともある)が結合すると考えてよい。
●現在では、エンハンサーという用語はあまり使わないことも多い。後述するが、エンハンサーにある、転写因子結合部位(シス認識配列)で議論をすることが多い。
●エンハンサーは、遺伝子からの距離、上流下流、エンハンサー配列の方向性に無関係に転写レベルを上昇させる。
●テストでどの用語を使おうか迷ったときは、今持っている教科書の用語を使えば良い。
●開始コドンより前、終止コドンより後ろに、非翻訳領域(翻訳されない領域)があることに注意せよ。転写開始点はプロモーターにあり、転写終結配列はターミネーターと呼ばれる。開始コドンや終止コドンは翻訳の開始と終止の合図であり、転写開始と転写終結の合図は別にあることに注意せよ。開始コドンから転写が始まるわけではない。開始コドンよりも前から転写が始まるのである。
●RNAは3’が伸びていく。3’にしかヌクレオチドを付加できないからである。
●リボソームはmRNA上を5‘から3’へ動いていく。
● 真核生物において、RNAポリメラーゼの結合するプロモーターの多くは、基本転写因子の結合するTATAボックスを含むが、TATAボックスを含まないTATAレスプロモーターも存在する。
●転写調節領域は、遺伝子調節領域とよばれることもある。遺伝子調節領域にはプロモーターを含むことがある。
●リプレッサーやアクチベーターの用語を真核生物の議論でも使うことがある。しかし、これらの用語は原核生物の議論で使われることが多い。
●エンハンサーには多数の活性化タンパク質が結合する領域がある。つまり、エンハンサーとは、近接した多数の活性化タンパク質結合部位群のことである。
エンハンサーにたくさんある活性化タンパク質結合部位のうち一つを指して、シス認識配列と呼ぶことがある。シスというのは同じ側という意味で、遺伝子と同じ染色体上にある調節配列なので、シス認識配列という。
●エンハンサーに活性化タンパク質と抑制タンパク質が競合的に結合し合うとする場合もある。つまりエンハンサーのみで議論し
、サイレンサーという概念を使わない場合もある(キャンベル生物学9版は、エンハンサーに活性化因子、抑制因子が結合するとしており、エンハンサーだけで議論をすすめている)。
活性化因子と抑制因子がDNAに結合する際に、部分的に同じ領域使う場合がある(2つの因子は同じ領域を取り合い、競合する)。つまり、エンハンサーとサイレンサーの違いはあいまいで、厳密な区別はできない。
●遺伝子から遠く離れた位置にないとエンハンサーやサイレンサーと呼ばないことが多い。したがって、エンハンサーやサイレンサーは、ふつう原核生物に使わない用語である。原核生物のオペレーターをサイレンサーとはふつう言わない。
● 真核生物の多くのmRNAの5' 末端にある修飾構造をキャップという。
必ずしも構造上の画一性はないが、5' 末端は必ず7-メチルグアノシンであり、その5' 部位と次のヌクレオシドの5' 部位が、三リン酸を介して結合している。
● 真核生物のmRNAの3' 末端に普遍的に存在するアデニル酸の連続した配列をポリAテール、もしくはポリA配列という。
その長さは数十から200ヌクレオチド程度であり、一つのmRNA分子種についても長さは一定でない。
ポリAテールは遺伝子によってコードされておらず、mRNA前駆体中のAAUAAAまたは類似の配列(ポリA配列付加シグナル)の3' 側へ10~30ヌクレオチドの位置で前駆体が切断された後、その3' 末端に付加される。この切断およびポリA付加反応(ポリアデニル化polyadenylation)は、核内で起こり、ポリAポリメラーゼ(poly-A polymerase)や、AAUAAA配列およびその3' 側の未確定のシグナル配列を必要とする。ポリA配列の機能は確定的でないが、mRNAの安定化、翻訳、核から細胞質への輸送、mRNAスプライシングの際の3'末端にあるエクソンのシグナルとしての役割などが示唆されている。
●複数の転写因子がひとつの遺伝子を制御するのが普通。
●転写因子の結合から必ず転写がスタートするとは限らない。ただ、複数の転写活性化因子が掛け算的に転写装置とDNAの親和性を高めていく(転写開始構造をとるためのエネルギーの障壁を低くしていく)のは間違いない。
●数千塩基対も離れた位置にあるエンハンサーがプロモーターにおける転写開始に影響を与える仕組みについては、完全には解明されていない。今回は、キャンベル生物学に従って、DNA湾曲タンパク質(DNAを折り曲げるタンパク質)が働いている仕組みを紹介した(また、板書していないが、転写因子とプロモーター付近のタンパク質を結び付けるタンパク質、すなわち仲介役のメディエータータンパク質が存在することは間違いない)。
このほか、コヒーシンと言う輪のようなタンパク質が折れ曲がったDNAを固定しているという説も有力である(トングを輪っかで固定するように、折れ曲がったDNAをコヒーシンが固定する。コヒーシンは資料集で見たことがあるだろう。コヒーシンの姉妹染色分体を固定する働きは有名である)。
*16:05「編集」は狭義のRNA editingを指していません。正確を期して「修飾」と言うべきでした。すみません。
0:00 転写調節の概要(タンパク質がDNAに結合する)
4:55 RNAポリメラーゼがプロモーターに結合する
7:22 真核生物の転写(転写因子と基本転写因子)
15:18 真核生物のRNAの修飾(プロセッシング)
19:24 余談(転写因子の存在パターン)
#高校生物
#分子生物学
#遺伝子
この動画を含むファイル
関連動画




関連用語
